Introduction
Hawthorn appears to function as a system modulator. Its effects extend across circulation, inflammation, pain signaling, central nervous system activity, and protection during tissue injury. These physiological domains are deeply interconnected within a living organism. Changes in one system influence the others through overlapping networks of signaling molecules, regulatory pathways, and physiological feedback loops.
The body operates through interacting regulatory systems that continuously shape one another’s behavior. A change in vascular function can alter inflammatory activity. Inflammation can affect pain sensitivity and neural excitability. Neural signaling can influence immune function and tissue repair. The resulting effects are often larger and more complex than would be predicted by examining any one system in isolation.
Contents
- Introduction
- Activity Across Physiological Systems
- Circulation
- Inflammation
- Pain
- CNS – Central Nervous System
- Injury Protection
- The Extract Matrix Effect
- From Components to Systems
- Candidate Unifying Mechanisms
- Redox Regulation and the Nrf2/HO-1 Pathway
- NF-κB and the Regulation of Inflammation
- Opioid Receptor Modulation
- Convergence, Crosstalk, and a Working Model
- Strengths, Limitations, and Research Gaps
- References
This perspective provides a useful framework for understanding hawthorn. Hawthorn affects several physiological domains that at first appear unrelated. At the same time, many of the same signaling pathways recur across those domains. The Nrf2/HO-1 stress-response system, NF-κB inflammatory signaling, opioid pathways, and related regulatory networks appear repeatedly in the literature. The pattern suggests that at least part of hawthorn’s apparent breadth may arise from effects on a limited number of interconnected regulatory hubs rather than from separate mechanisms for every observed outcome.
The situation is further complicated by the nature of botanical extracts themselves. Hawthorn is not a single compound but a complex mixture of flavonoids, procyanidins, organic acids, and other constituents that interact with one another. Whole extracts behave differently from isolated compounds, producing pharmacokinetic and biological effects that cannot be predicted from their individual components. Interactions among constituents can amplify, extend, or reshape activity in ways that are difficult to capture through reductionist models.
This article develops a working mechanistic picture of hawthorn as a system modulator. It begins by reviewing the major domains in which hawthorn activity has been documented. It then examines the pharmacokinetic behavior of whole extracts and the candidate mechanisms that appear most consistently across the literature. A relatively small number of interacting processes may give rise to the broad physiological profile repeatedly observed with hawthorn extracts.
Activity Across Physiological Systems
Hawthorn’s effects are clear in several physiological systems. The categories described below are organizational rather than biological. In the living organism these systems continuously interact, but examining them separately makes it easier to see the patterns before turning to the mechanisms that may connect them.
Circulation
This is the domain with the most substantial body of evidence. Hawthorn extracts have been shown in multiple studies to support endothelial function, promote nitric-oxide-mediated vasodilation, exert mild cardiotonic effects, and slow aspects of atherosclerotic progression through combined lipid-lowering, antioxidant, and anti-inflammatory actions. These effects are not attributed to a single compound. Flavonoids and oligomeric procyanidins appear to contribute to vessel-wall stability and capillary integrity, a pattern also seen with related polyphenols in other vascular contexts.
Human data are strongest here. Standardized extracts have been studied in people with mild heart failure and in those with elevated cardiovascular risk markers, with improvements reported in exercise tolerance, symptoms, and certain hemodynamic measures. Preclinical work has identified several plausible mechanisms, including protection of endothelial cells under stress, enhancement of nitric oxide bioavailability, and reduction of oxidative lipid modification involved in plaque formation.
A related area of interest is small-vessel and capillary health. Traditional use and modern research both point toward improved resilience of fine vessels. Informal observations across years of use demonstrated sustained normalization of previously elevated blood pressure together with a marked reduction in spontaneous bruising. The reduction in bruising was more reliable than that observed with sassafras tincture, which had been used for the same purpose. While these observations remain anecdotal, they align with the documented effects on endothelial function and capillary integrity.
Inflammation
Inflammation is a recurring theme in hawthorn research. Animal and cellular studies have repeatedly demonstrated anti-inflammatory effects from a variety of hawthorn extracts and constituents. Reported findings include reductions in pro-inflammatory cytokines and improvements in tissues exposed to inflammatory stress.
Several studies have documented decreases in inflammatory markers including TNF-α, IL-1β, and IL-6 in models of vascular, metabolic, and hepatic stress. These effects have been observed across multiple preparations and experimental systems, suggesting that modulation of inflammatory activity is a consistent feature of hawthorn’s physiological profile.
The picture is not entirely uniform. Some polysaccharide-rich extracts have produced immune-activating effects in isolated cell systems, indicating that responses can vary with preparation, concentration, and biological context. This variability is common in botanical research and highlights the importance of distinguishing between different hawthorn species, plant parts, and extraction methods.
Taken as a whole, the literature points toward a predominantly anti-inflammatory effect in living systems experiencing chronic low-grade inflammatory or oxidative stress.
Pain
Pain is one of the more intriguing areas of hawthorn research because the evidence points toward a specific functional mechanism rather than only broad antioxidant or anti-inflammatory effects. A 2010 study in mice examined hawthorn pulp and seed extracts across three standard pain models: mechanical (tail-clip), thermal (hot-plate), and chemical (acetic acid-induced writhing). Both extracts produced dose-dependent analgesia. Pretreatment with the opioid receptor antagonist naloxone completely abolished these effects in all three tests. The same extracts also reduced spontaneous locomotor activity and exploratory behavior, consistent with a calming or sedative component alongside pain relief.
These findings strongly implicate the endogenous opioid system. The naloxone reversal indicates that the analgesic effect depends on opioid receptor activity, either through direct effects of extract constituents or through facilitation of the body’s own opioid signaling. Follow-up analysis suggested greater involvement of kappa and delta receptor subtypes than mu receptors, based on the pattern of responses observed.
Additional evidence comes from studies of individual hawthorn constituents. Two flavonoids found in hawthorn, hyperoside and apigenin, have demonstrated opioid-receptor activity in binding and functional studies. Hyperoside showed preference for kappa receptors with relatively high delta affinity, while apigenin displayed a more balanced profile across mu, delta, and kappa receptors. The contribution of these compounds to the effects of whole extracts remains uncertain.
One notable observation is the difference between whole-extract behavior and isolated-compound studies. Hawthorn extracts produce clear naloxone-reversible analgesia, while some individual flavonoids display more complex receptor profiles when studied in isolation. This illustrates a recurring theme in botanical pharmacology: the behavior of a whole extract cannot always be predicted from the properties of its individual constituents.
CNS – Central Nervous System
Effects on the central nervous system appear in both traditional use and modern research, primarily in the direction of reduced excitability and improved calm. Sedative and anxiolytic actions have been documented in animal models, with several mechanisms proposed to contribute to these effects.
A 2022 study of Crataegus pinnatifida found clear anxiolytic and antidepressant-like effects in stressed mice. These effects were associated with activation of the 5-HT1A receptor and increased levels of brain-derived neurotrophic factor (BDNF) in the hippocampus and prefrontal cortex. The behavioral improvements were comparable in magnitude to those produced by a standard antidepressant in the same model.
Hawthorn extracts and several of their constituents have also been shown to inhibit acetylcholinesterase, the enzyme responsible for breaking down acetylcholine. This effect has been observed across multiple Crataegus species and is one proposed route by which hawthorn may support memory and learning. In animal models of induced memory impairment, improvements in behavioral testing have been accompanied by reductions in acetylcholinesterase activity.
Taken together, these findings point to genuine neuromodulatory activity rather than an indirect byproduct of vascular, inflammatory, or metabolic changes elsewhere in the organism. Multiple signaling systems appear to be involved, including serotonergic pathways, cholinergic pathways, and other mechanisms associated with regulation of excitability, mood, and cognitive function.
Injury Protection
Hawthorn extracts have been studied in models of acute ischemia-reperfusion injury in both the brain and heart. These injuries occur when blood flow is temporarily interrupted and then restored, triggering a surge of oxidative stress, inflammation, and secondary tissue damage. Across multiple studies, hawthorn treatment has been associated with reduced injury severity and improved functional outcomes.
Reported benefits include smaller areas of tissue death (reduced infarct volume), lower levels of oxidative stress markers, preservation of vulnerable tissue, and improved neurological or cardiac function following injury. These findings have been observed with both whole extracts and isolated hawthorn constituents.
Follow-up work examining the same stroke model identified an immunomodulatory component to this protection. Hawthorn pretreatment increased the presence of regulatory T cells and IL-10 in injured brain tissue, alongside elevated STAT-3 phosphorylation associated with reduced apoptosis during reperfusion. This pattern points to active suppression of the post-injury inflammatory cascade, a distinct mechanism from antioxidant protection alone, and one that connects directly to the broader inflammatory regulation discussed later in this article.
The protective effects are particularly notable because they appear in tissues that differ substantially in structure and function. Similar patterns have been reported in both neural and cardiovascular injury models, suggesting that hawthorn is acting on fundamental stress-response and tissue-protection pathways rather than on mechanisms unique to a single organ system.
Taken together, these findings indicate a consistent capacity to limit tissue damage and support recovery during acute physiological stress. The underlying mechanisms are explored later in this article.
The Extract Matrix Effect
One practical consideration shapes how all of the preceding findings are interpreted: hawthorn is administered as a complex extract, not as a single isolated molecule. Once in the body, the constituents of that extract do not behave independently. They influence one another’s absorption, distribution, metabolism, and persistence in circulation.
As a result, constituents delivered as part of a whole extract often reach higher blood concentrations, remain in the system longer, and produce greater overall exposure than the same compounds administered in purified form. This phenomenon, often called the matrix effect, is well recognized in the study of complex botanical preparations.
The extract contains flavonoids, procyanidins, organic acids, and numerous other compounds. Together they can alter solubility, transport, metabolism, and clearance in ways that increase or prolong biological activity. The result is that the activity of the extract is frequently greater or more sustained than would be predicted from any single constituent studied in isolation.
This observation has practical implications. It helps explain why attempts to identify a single “active ingredient” often fail to capture the behavior of the whole preparation. It also helps explain why standardized extracts frequently outperform highly purified fractions despite containing many compounds at lower concentrations than would be used in isolation.
More importantly, the matrix effect introduces a broader principle. When multiple constituents with overlapping or complementary actions remain present together at meaningful levels, they can influence numerous pathways simultaneously. The resulting biological activity reflects not only the properties of the individual compounds but also their interactions. The behavior of the whole extract emerges from the relationships among its parts.
From Components to Systems
The same pattern appears repeatedly throughout biology. Individual molecules participate in pathways. Pathways interact with one another through regulatory networks. Those networks influence physiological systems that continuously exchange information and adapt to changing conditions. At each level, the behavior of the whole is shaped by interactions among its components.
This perspective provides a framework for understanding hawthorn. The preceding sections described activity in circulation, inflammation, pain, central nervous system function, and protection during acute injury. These observations were presented separately for clarity, but the underlying biology is not organized into separate compartments. The same signaling pathways often participate in multiple physiological processes, linking systems that are traditionally studied in isolation.
The matrix effect demonstrates that the activity of a botanical extract cannot always be predicted from the behavior of its individual constituents. A similar principle may apply at higher levels of organization. The breadth of hawthorn’s physiological effects may arise not from a different mechanism for every observation, but from influence on a relatively small number of regulatory processes that participate in many systems simultaneously.
This does not imply that a single pathway explains everything. Biological regulation is too complex for that. It does suggest that the search for a handful of recurring mechanisms may be more productive than treating each observed effect as an entirely separate phenomenon.
With that framework in place, we can now examine the candidate mechanisms that appear most consistently across the literature.
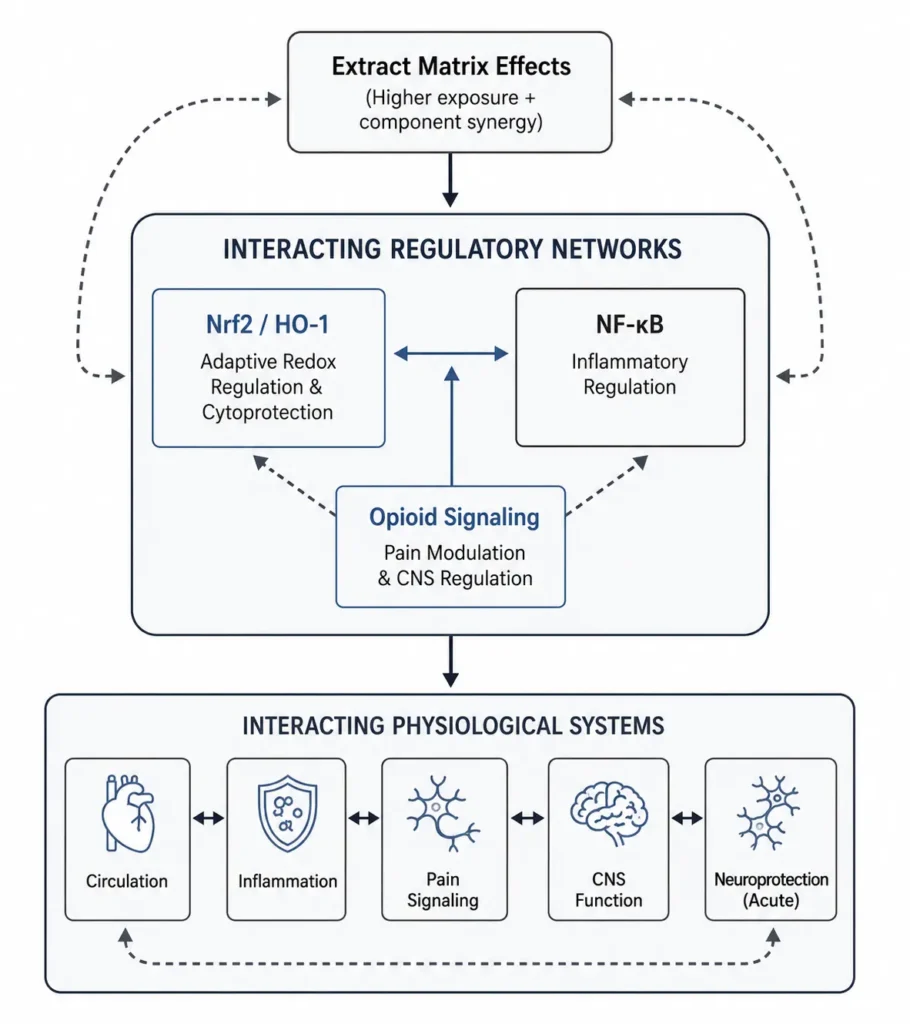
A system modulator resonates through interacting networks whose effects fold and refold through the biology of the organism as it adapts and responds.
Candidate Unifying Mechanisms
Several recurring mechanisms appear throughout the hawthorn literature. Among them, four stand out because they connect multiple physiological systems and repeatedly emerge across otherwise separate lines of research: redox regulation through the Nrf2/HO-1 pathway, modulation of NF-κB signaling, engagement of opioid pathways, and the extensive crosstalk among these processes.
The sections that follow examine each of these mechanisms and their potential contribution to the broader pattern of effects observed across circulation, inflammation, pain, central nervous system function, and protection during acute injury.
Redox Regulation and the Nrf2/HO-1 Pathway
Nrf2 is one of the body’s primary adaptive-response systems. Cells continuously monitor their internal environment for signs of oxidative, metabolic, or chemical stress. When stress rises, Nrf2 moves into the nucleus and activates a coordinated program of protective genes that help restore cellular balance and resilience.
One of the most important outputs of this response is heme oxygenase-1 (HO-1). HO-1 breaks down heme into several biologically active products, including carbon monoxide and biliverdin. Carbon monoxide can promote vasodilation and help moderate inflammatory signaling, while biliverdin and its downstream product bilirubin contribute antioxidant and cytoprotective effects. Together, these responses support vascular function, cellular survival, and recovery from stress.
Hawthorn extracts and several of their flavonoid constituents have been shown to activate this pathway. In one study, hawthorn fruit extract increased Nrf2 and HO-1 expression in an animal model while improving lipid profiles. Other work with Crataegus oxyacantha bark extracts demonstrated Nrf2 nuclear translocation and induction of HO-1 along with other protective enzymes. A hawthorn-derived flavonoid has also been linked to dual activation of AMPK and Nrf2 with improvements in mitochondrial function under stress.
These findings provide direct mechanistic support for many of the observations reviewed earlier. Activation of the Nrf2/HO-1 axis can strengthen endothelial resilience, improve vascular function, reduce oxidative damage, and increase resistance to acute physiological stress. Because Nrf2 also interacts extensively with inflammatory signaling networks, its influence extends beyond redox regulation alone.
Among the mechanisms discussed in this review, Nrf2/HO-1 stands out as a particularly plausible regulatory hub. Its effects span multiple physiological systems, and its activation provides a coherent explanation for a substantial portion of the circulatory, anti-inflammatory, and tissue-protective activity observed with hawthorn extracts.
NF-κB and the Regulation of Inflammation
NF-κB is one of the body’s primary inflammatory regulators. When activated by infection, tissue injury, cytokines, or other stress signals, it enters the nucleus and drives expression of genes involved in inflammation, immune-cell recruitment, and tissue defense. These include cytokines such as TNF-α, IL-1β, and IL-6, along with numerous chemokines, adhesion molecules, and inflammatory enzymes.
Hawthorn extracts and their constituents have repeatedly been shown to reduce NF-κB activation or suppress expression of NF-κB-dependent inflammatory genes. These findings align with the reductions in inflammatory cytokines observed across a variety of vascular, metabolic, and tissue-injury models.
The most interesting aspect of NF-κB biology, however, is its relationship with Nrf2. These pathways participate in a continuous regulatory dialogue that helps determine whether stress responses become self-limited and adaptive or escalate into persistent inflammation and tissue damage.
Activation of Nrf2 generally restrains NF-κB signaling through several complementary mechanisms. Increased antioxidant capacity reduces the oxidative conditions that favor NF-κB activation. Nrf2 signaling can stabilize inhibitory proteins that keep NF-κB inactive, and the two pathways compete for some of the same transcriptional resources within the nucleus. The overall effect is a shift away from inflammatory amplification and toward restoration of cellular homeostasis.
The relationship is not strictly one-way. NF-κB activity can, under some conditions, contribute to activation of Nrf2-dependent defenses as part of a feedback response to inflammation and oxidative stress. Health depends not on elimination of either pathway but on appropriate balance between them. Acute NF-κB activation helps contain threats and coordinate repair, while Nrf2 promotes recovery, resolution, and restoration of cellular function.
Viewed in this context, hawthorn’s anti-inflammatory effects are not simply a matter of suppressing inflammatory mediators. They may reflect influence on a broader regulatory system that continuously balances defense, adaptation, repair, and recovery. This interaction between Nrf2 and NF-κB provides one of the strongest mechanistic bridges connecting the circulatory, inflammatory, and tissue-protective effects reviewed earlier.
Opioid Receptor Modulation
The strongest evidence for opioid-system involvement comes from a 2010 study in which hawthorn pulp and seed extracts produced dose-dependent analgesia across mechanical, thermal, and chemical pain models in mice. Pretreatment with the opioid antagonist naloxone abolished these effects in every model tested, providing direct functional evidence that opioid pathways were required for the observed analgesia.
The same extracts also reduced spontaneous locomotor activity and exploratory behavior, suggesting effects that extended beyond pain perception alone. Together, these findings point toward a broader influence on neural excitability and behavioral state.
Additional evidence comes from studies of individual hawthorn constituents. Two flavonoids present in hawthorn, hyperoside and apigenin, have demonstrated opioid-receptor activity in receptor-binding and functional studies. Hyperoside showed preference for kappa receptors with relatively high delta affinity, while apigenin displayed a more balanced profile across mu, delta, and kappa receptors. The contribution of these compounds to the activity of whole extracts remains uncertain.
As with other areas of hawthorn research, the behavior of the whole extract appears more straightforward than the behavior of its individual constituents. Hawthorn extracts produce clear naloxone-sensitive analgesia, while isolated compounds display more complex and variable receptor profiles. This pattern is consistent with the broader theme of matrix effects and coordinated activity across multiple constituents.
Within the working model developed here, opioid signaling offers a direct explanation for many of the pain-relief and CNS-calming observations reviewed earlier. It may also participate in wider stress-response networks. Endogenous opioid systems influence not only pain perception but also emotional regulation, stress resilience, and aspects of tissue response to injury. Whether hawthorn engages these broader functions remains incompletely understood, but the possibility provides a plausible point of connection between pain modulation, reduced excitability, and protection during physiological stress.
Convergence, Crosstalk, and a Working Model
A coherent picture begins to emerge when these mechanisms are viewed as parts of an interacting system rather than as independent explanations. Nrf2, NF-κB, and opioid signaling each influence different aspects of physiology, but none operates in isolation. They are components of overlapping regulatory networks that respond to stress, injury, inflammation, and adaptation.
At the pathway level, Nrf2 and NF-κB form a particularly important regulatory pair. Nrf2 promotes cellular protection, repair, and recovery, while NF-κB coordinates inflammatory responses to threat and injury. Their continuous interaction helps determine whether a challenge resolves through adaptation and restoration or progresses toward persistent inflammation and tissue damage. Hawthorn’s ability to influence both sides of this relationship provides a plausible explanation for the combination of circulatory support, anti-inflammatory effects, and protection during acute physiological stress observed throughout the literature.
Opioid signaling appears to participate in a partially overlapping network. Its most direct contributions are seen in pain modulation, reduced excitability, and shifts toward behavioral calm. At the same time, endogenous opioid systems are deeply integrated with broader stress-response pathways. This raises the possibility that opioid signaling contributes to a wider pattern of physiological regulation than analgesia alone, although the extent of that contribution remains incompletely understood.
The extract matrix effect adds another layer to the picture. Hawthorn is not a single active molecule acting through a single target. It is a complex preparation whose constituents alter one another’s absorption, persistence, and biological activity. Multiple compounds remain present simultaneously, influencing multiple pathways at the same time. The resulting effects are shaped not only by the actions of individual constituents but also by their interactions.
The same organizational principle appears at every level examined in this review. Constituents interact within the extract. Signaling pathways interact within regulatory networks. Physiological systems interact within the organism. The behavior of the whole cannot always be predicted from the behavior of the isolated parts.
This perspective helps explain why hawthorn repeatedly appears in physiological domains that initially seem unrelated. Circulation, inflammation, pain signaling, central nervous system activity, and protection during acute injury are often studied as separate subjects, yet they are connected through overlapping networks of regulation and response. Influencing a small number of strategic regulatory hubs can therefore produce effects that propagate across multiple systems simultaneously.
This remains a working model rather than a settled mechanism. It does not require every effect to travel through the same pathway, nor does it exclude additional contributors such as nitric oxide signaling, AMPK activation, mitochondrial effects, or mechanisms that have not yet been identified. What it offers is a framework for understanding how a single botanical preparation can produce a broad and coherent physiological profile without requiring a separate explanation for every observation.
Viewed through this lens, hawthorn appears less like a collection of isolated activities and more like a system modulator: a botanical that influences interacting regulatory networks whose effects extend across the integrated physiology of the organism.
Strengths, Limitations, and Research Gaps
Several aspects of this model are supported by substantial evidence. Cardiovascular effects have been studied extensively in both experimental and human settings. Anti-inflammatory activity has been demonstrated across multiple preparations and biological systems. Opioid-dependent analgesia has direct functional support, and activation of the Nrf2/HO-1 pathway has been documented in several hawthorn-derived preparations.
Other aspects remain more tentative. Much of the mechanistic literature is preclinical, and relatively few studies measure multiple pathways simultaneously. The proposed relationships among Nrf2, NF-κB, opioid signaling, and the observed physiological effects are biologically plausible and supported by existing evidence, but they have rarely been examined together within the same experimental framework.
The extract matrix effect presents an additional challenge. Whole extracts often behave differently than isolated constituents, making it difficult to assign observed outcomes to individual compounds. This complexity is part of what makes botanical preparations scientifically interesting, but it also complicates mechanistic interpretation.
Research on acute injury models is more extensive than research on chronic injury and recovery. Whether the same mechanisms contribute to long-term resilience, adaptation, or repair remains an important question for future study.
The model presented here should therefore be viewed as a working framework rather than a settled conclusion. Its value lies in organizing a large and diverse body of observations into a coherent picture that can be tested, refined, or revised as additional evidence emerges.
A related (and more readable) article appears here: https://vitalsystems.substack.com/p/hawthorn-a-system-modulator
References
1. Can ÖD, Özkay ÜD, Öztürk N, Öztürk Y. Effects of hawthorn seed and pulp extracts on the central nervous system in mice. Pharmaceutical Biology. 2010;48(8):924-931. doi:10.3109/13880200903305500
https://doi.org/10.3109/13880200903305500
2. Yoo JH, Liu Y, Kim HS. Hawthorn fruit extract elevates expression of Nrf2/HO-1 and improves lipid profiles in ovariectomized rats. Journal of Medicinal Food. 2016;19(7):643-650. doi:10.1089/jmf.2015.3600
https://doi.org/10.1089/jmf.2015.3600
3. Nitzan K, et al. Anxiolytic and antidepressants’ effect of Crataegus pinnatifida (Shan Zha): biochemical mechanisms. Translational Psychiatry. 2022;12:204. doi:10.1038/s41398-022-01970-6
https://doi.org/10.1038/s41398-022-01970-6
4. Bahorun T, et al. Hawthorn (Crataegus oxyacantha L.) bark extract regulates antioxidant response element (ARE)-mediated enzyme expression via Nrf2 pathway activation in normal hepatocyte cell line. Phytotherapy Research. 2015;29(11):1743-1752. doi:10.1002/ptr.5035
https://doi.org/10.1002/ptr.5035
5. Katavic PL, et al. Flavonoids as opioid receptor ligands: identification and preliminary structure-activity relationships. Journal of Natural Products. 2007;70(8):1275-1280. doi:10.1021/np070194x
https://doi.org/10.1021/np070194x
6. Wu M, et al. Roles and mechanisms of hawthorn and its extracts on cardiovascular and cerebrovascular diseases. Frontiers in Pharmacology. 2020;11:118. doi:10.3389/fphar.2020.00118
https://doi.org/10.3389/fphar.2020.00118
7. Elango C, et al. Immunomodulatory effect of hawthorn extract in an experimental stroke model. Journal of Neuroinflammation. 2010;7:97. doi:10.1186/1742-2094-7-97
https://doi.org/10.1186/1742-2094-7-97